
The exact mechanisms underlying these diseases are still not fully understood but increasing evidence suggests that dietinduced monocytes and macrophage dysfunction plays a major causal role in the conversion of acute into chronic inflammation and ID: 931316
Download Presentation The PPT/PDF document "Atherosclerosis is a progressive chronic..." is the property of its rightful owner. Permission is granted to download and print the materials on this web site for personal, non-commercial use only, and to display it on your personal computer provided you do not modify the materials and that you retain all copyright notices contained in the materials. By downloading content from our website, you accept the terms of this agreement.













Slide1
Atherosclerosis is a progressive chronic inflammatory disease and the main contributor to cardiovascular diseases (CVD) worldwide [1, 2].
The exact mechanisms underlying these diseases are still not fully understood, but increasing evidence suggests that diet-induced monocytes and macrophage dysfunction plays a major causal role in the conversion of acute into chronic inflammation and
atherogenesis
[3, 4]. Current lipid-lowering therapies have only a limited effect on CVD mortality [5], particularly in patients unable to tolerate statins (15-20%) [6]. Metabolic stress enhances monocyte infiltration into atherosclerotic plaques by rendering blood monocytes hyper-chemotactic and dysfunctional through a process that involves increased oxidative stress and is mediated by protein S-glutathionylation [7, 8].Monocyte priming and reprogramming induced by metabolic stress is prevented by overexpression of glutaredoxin 1 (Grx1) [9, 10].Epigenetic enzymes regulate monocyte and macrophage functions, and targeting epigenetic enzymes has proven to be an effective tool to dampen inflammatory responses [11, 12]. Histone deacetylases (HDACs) are part of a large family of enzymes that play crucial roles in numerous biological processes, largely through their repressive effects on transcription [13, 14]. They mediate lysine deacetylation, regulate immunometabolism and inflammatory outputs from macrophages [15].
Inhibition of Myeloid HDAC2 Upregulates
Glutaredoxin
1 Expression Improves Protein Thiol Redox State and Protects against High-Calorie Diet-Induced Monocyte Dysfunction and Atherosclerosis.
Methodology
Results
Introduction
Hypothesis
Conclusion
Reference
1. Galkina, E. and K. Ley, Immune and inflammatory mechanisms of atherosclerosis (*). Annual review of immunology, 2009. 27: p. 165-197.2. Manduteanu, I. and M. Simionescu, Inflammation in atherosclerosis: a cause or a result of vascular disorders? J Cell Mol Med, 2012. 16(9): p. 1978-90.3. Groh, L., et al., Monocyte and macrophage immunometabolism in atherosclerosis. 2018. 40(2): p. 203-214.4. Moore, K.J., F.J. Sheedy, and E.A. Fisher, Macrophages in atherosclerosis: a dynamic balance. Nature reviews. Immunology, 2013. 13(10): p. 709-721.5. Butters, J., et al., Clinical Outcomes in Trials Evaluating Lipid-Lowering Drugs. Am J Cardiovasc Drugs., 2017. 17(6): p. 447-452.6. Ward, N.C., G.F. Watts, and R.H. Eckel, Statin Toxicity. 2019. 124(2): p. 328-350.7. Kim, H.S., et al., Redox regulation of MAPK phosphatase 1 controls monocyte migration and macrophage recruitment. Proceedings of the National Academy of Sciences, 2012. 109(41): p. 16422-16423.8. Ullevig, S.L., et al., Protein S-Glutathionylation Mediates Macrophage Responses to Metabolic Cues from the Extracellular Environment. Antioxid Redox Signal, 2016. 25(15): p. 836-851.9. Downs, K., et al., Monocytic Glutaredoxin 1 Protects Mice against Obesity, Hyperglycemia and Atherosclerosis. Free Radical Biology and Medicine, 2016. 100: p. S53.10. Ahn, Y.J., Wang, L., Tavakoli, S., Nguyen, H.N., Short, J. and Asmis, , Glutaredoxin 1 controls the sexual dimorphic reprogramming of monocytes by nutrient stress and protects mice against obesity, hyperglycemia and atherosclerosis. . 2019: In preparation for Cell Metabolism.11. A., H.M. and d.W.M. P.J., Epigenetic Regulation of Monocyte and Macrophage Function. Antioxidants & Redox Signaling, 2016. 25(14): p. 758-774.12. Shakespear, M.R., et al., Lysine Deacetylases and Regulated Glycolysis in Macrophages. Trends Immunol, 2018. 39(6): p. 473-488.13. Gallinari, P., et al., HDACs, histone deacetylation and gene transcription: from molecular biology to cancer therapeutics. Cell Research, 2007. 17(3): p. 195-211.14. Murray, P.J. and T.A. Wynn, Protective and pathogenic functions of macrophage subsets. Nature reviews. Immunology, 2011. 11(11): p. 723-737.15. Jeong, Y., et al., Histone deacetylase isoforms regulate innate immune responses by deacetylating mitogen-activated protein kinase phosphatase-1. J Leukoc Biol, 2014. 95(4): p. 651-9.16. Ahn, Y.J., Wang, L., and Asmis, R., Quantification of Monocyte Chemotactic Activity In Vivo and Characterization of Blood Monocyte Derived Macrophages. JoVE, 2019(150): p. e59706.17. Qiao, M., et al., Thiol oxidative stress induced by metabolic disorders amplifies macrophage chemotactic responses and accelerates atherogenesis and kidney injury in LDL receptor-deficient mice. Arteriosclerosis, thrombosis, and vascular biology, 2009. 29(11): p. 1779-1786.
Luxi Wang, Yong Joo Ahn and Reto Asmis
Wake Forest University School of Medicine
We hypothesize that by inducing Grx1, HDAC2 inhibition protects monocytes and macrophages from HCD-induced dysfunction by preventing nutrient stress-induced protein
S-glutathionylation, thereby preserving redox signaling pathways and cell function and restoring macrophage plasticity and inflammation-resolving activity.
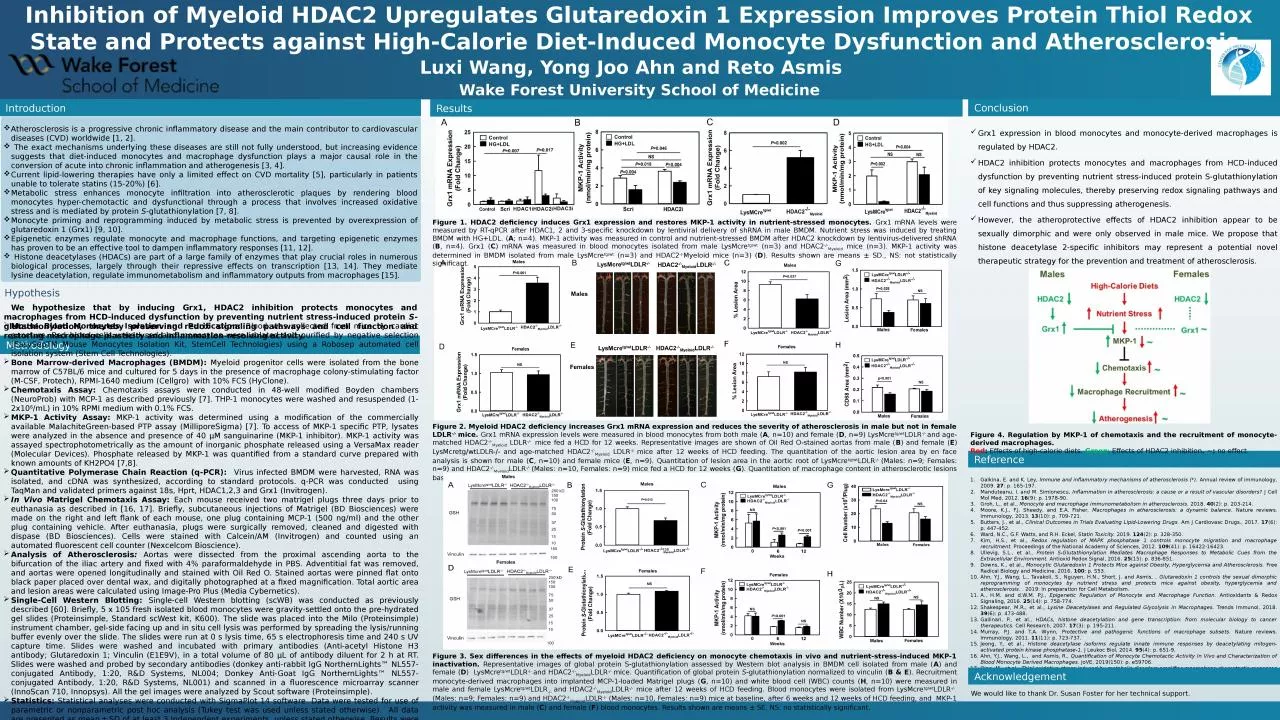
Grx1 expression in blood monocytes and monocyte-derived macrophages is regulated by HDAC2. HDAC2 inhibition protects monocytes and macrophages from HCD-induced dysfunction by preventing nutrient stress-induced protein S-glutathionylation of key signaling molecules, thereby preserving redox signaling pathways and cell functions and thus suppressing atherogenesis.However, the atheroprotective effects of HDAC2 inhibition appear to be sexually dimorphic and were only observed in male mice. We propose that histone deacetylase 2-specific inhibitors may represent a potential novel therapeutic strategy for the prevention and treatment of atherosclerosis.
Figure 1. HDAC2 deficiency induces Grx1 expression and restores MKP-1 activity in nutrient-stressed monocytes. Grx1 mRNA levels were measured by RT-qPCR after HDAC1, 2 and 3-specific knockdown by lentiviral delivery of shRNA in male BMDM. Nutrient stress was induced by treating BMDM with HG+LDL. (A; n=4). MKP-1 activity was measured in control and nutrient-stressed BMDM after HDAC2 knockdown by lentivirus-delivered shRNA (B, n=4). Grx1 (C) mRNA was measured in blood monocytes isolated from male LysMcretg/wt (n=3) and HDAC2-/-Myeloid mice (n=3). MKP-1 activity was determined in BMDM isolated from male LysMcretg/wt (n=3) and HDAC2-/-Myeloid mice (n=3) (D). Results shown are means ± SD., NS: not statistically significant.
Figure 3. Sex differences in the effects of myeloid HDAC2 deficiency on monocyte chemotaxis in vivo and nutrient-stress-induced MKP-1 inactivation. Representative images of global protein S-glutathionylation assessed by Western blot analysis in BMDM cell isolated from male (A) and female (D) LysMcretg/wtLDLR-/- and HDAC2-/-MyeloidLDLR-/- mice. Quantification of global protein S-glutathionylation normalized to vinculin (B & E). Recruitment monocyte-derived macrophages into implanted MCP-1-loaded Matrigel plugs (G, n=10) and white blood cell (WBC) counts (H, n=10) were measured in male and female LysMcretg/wtLDLR-/- and HDAC2-/-MyeloidLDLR-/- mice after 12 weeks of HCD feeding. Blood monocytes were isolated from LysMcretg/wtLDLR-/- (Males: n=9; Females: n=9) and HDAC2-/-MyeloidLDLR-/- (Males: n=10, Females: n=9) mice at baseline, after 6 weeks and 12 weeks of HCD feeding, and MKP-1 activity was measured in male (C) and female (F) blood monocytes. Results shown are means ± SE. NS: no statistically significant.
Figure 2. Myeloid HDAC2 deficiency increases Grx1 mRNA expression and reduces the severity of atherosclerosis in male but not in female LDLR-/- mice. Grx1 mRNA expression levels were measured in blood monocytes from both male (A, n=10) and female (D, n=9) LysMcretg/wtLDLR-/- and age-matched HDAC2-/-Myeloid LDLR-/- mice fed a HCD for 12 weeks. Representative images are shown of Oil Red O-stained aortas from male (B) and female (E) LysMcretg/wtLDLR-/- and age-matched HDAC2-/-Myeloid LDLR-/- mice after 12 weeks of HCD feeding. The quantitation of the aortic lesion area by en face analysis is shown for male (C, n=10) and female mice (E, n=9). Quantitation of lesion area in the aortic root of LysMcretg/wtLDLR-/- (Males: n=9; Females: n=9) and HDAC2-/-MyeloidLDLR-/- (Males: n=10, Females: n=9) mice fed a HCD for 12 weeks (G). Quantitation of macrophage content in atherosclerotic lesions based on CD68 staining in the aortic root (H). Results shown are means ± SE. NS: not statistically significant.
Mouse Blood Monocytes Isolation and Purification: Blood was collected from mice by cardiac puncture. Red blood cells were lysed and monocytes were isolated and purified by negative selection (EasySepTM Mouse Monocytes Isolation Kit, StemCell Technologies) using a Robosep automated cell isolation system (Stem Cell Technologies).Bone Marrow-derived Macrophages (BMDM): Myeloid progenitor cells were isolated from the bone marrow of C57BL/6 mice and cultured for 5 days in the presence of macrophage colony-stimulating factor (M-CSF, Protech), RPMI-1640 medium (Cellgro) with 10% FCS (HyClone). Chemotaxis Assay: Chemotaxis assays were conducted in 48-well modified Boyden chambers (NeuroProb) with MCP-1 as described previously [7]. THP-1 monocytes were washed and resuspended (1-2x106/mL) in 10% RPMI medium with 0.1% FCS.MKP-1 Activity Assay: MKP-1 activity was determined using a modification of the commercially available MalachiteGreen-based PTP assay (MilliporeSigma) [7]. To access of MKP-1 specific PTP, lysates were analyzed in the absence and presence of 40 μM sanguinarine (MKP-1 inhibitor). MKP-1 activity was assayed spectrophotometrically as the amount of inorganic phosphate released using a VersaMax reader (Molecular Devices). Phosphate released by MKP-1 was quantified from a standard curve prepared with known amounts of KH2PO4 [7,8]. Quantitative Polymerase Chain Reaction (q-PCR): Virus infected BMDM were harvested, RNA was isolated, and cDNA was synthesized, according to standard protocols. q-PCR was conducted using TaqMan and validated primers against 18s, Hprt, HDAC1,2,3 and Grx1 (Invitrogen). In Vivo Matrigel Chemotaxis Assay: Each mouse received two matrigel plugs three days prior to euthanasia as described in [16, 17]. Briefly, subcutaneous injections of Matrigel (BD Biosciences) were made on the right and left flank of each mouse, one plug containing MCP-1 (500 ng/ml) and the other plug containing vehicle. After euthanasia, plugs were surgically removed, cleaned and digested with dispase (BD Biosciences). Cells were stained with Calcein/AM (Invitrogen) and counted using an automated fluorescent cell counter (Nexcelcom Bioscience).Analysis of Atherosclerosis: Aortas were dissected from the proximal ascending aorta to the bifurcation of the iliac artery and fixed with 4% paraformaldehyde in PBS. Adventitial fat was removed, and aortas were opened longitudinally and stained with Oil Red O. Stained aortas were pinned flat onto black paper placed over dental wax, and digitally photographed at a fixed magnification. Total aortic area and lesion areas were calculated using Image-Pro Plus (Media Cybernetics). Single-Cell Western Blotting: Single-cell Western blotting (scWB) was conducted as previously described [60]. Briefly, 5 x 105 fresh isolated blood monocytes were gravity-settled onto the pre-hydrated gel slides (Proteinsimple, Standard scWest kit, K600). The slide was placed into the Milo (Proteinsimple) instrument chamber, gel-side facing up and in situ cell lysis was performed by spreading the lysis/running buffer evenly over the slide. The slides were run at 0 s lysis time, 65 s electrophoresis time and 240 s UV capture time. Slides were washed and incubated with primary antibodies (Anti-acetyl Histone H3 antibody; Glutaredoxin 1; Vinculin (E1E9V), in a total volume of 80 µL of antibody diluent for 2 h at RT. Slides were washed and probed by secondary antibodies (donkey anti-rabbit IgG NorthernLights™ NL557-conjugated Antibody, 1:20, R&D Systems, NL004; Donkey Anti-Goat IgG NorthernLights™ NL557-conjugated Antibody, 1:20, R&D Systems, NL001) and scanned in a fluorescence microarray scanner (InnoScan 710, Innopsys). All the gel images were analyzed by Scout software (Proteinsimple).Statistics: Statistical analyses were conducted with SigmaPlot 14 software. Data were tested for use of parametric or nonparametric post hoc analysis (Tukey test was used unless stated otherwise). All data are presented as mean ± SD of at least 3 independent experiments, unless stated otherwise. Results were considered statistically significant at the P < 0.05 level.
Acknowledgement
We would like to thank Dr. Susan Foster for her technical support.
Figure 4. Regulation by MKP-1 of chemotaxis and the recruitment of monocyte-derived macrophages. Red: Effects of high-calorie diets. Green: Effects of HDAC2 inhibition. ~: no effect.