PDF-Improvement of tropane alkaloids production in hair y
Author : test | Published Date : 2015-05-05
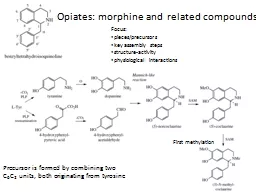
educn or zhihualiao163com Abstract Atropa belladonna L is the commercial plant to produce tropane alka loids including hyoscyamine and scopolamine which are widely
Presentation Embed Code
Download Presentation
Download Presentation The PPT/PDF document "Improvement of tropane alkaloids product..." is the property of its rightful owner. Permission is granted to download and print the materials on this website for personal, non-commercial use only, and to display it on your personal computer provided you do not modify the materials and that you retain all copyright notices contained in the materials. By downloading content from our website, you accept the terms of this agreement.
Improvement of tropane alkaloids production in hair y: Transcript
Download Rules Of Document
"Improvement of tropane alkaloids production in hair y"The content belongs to its owner. You may download and print it for personal use, without modification, and keep all copyright notices. By downloading, you agree to these terms.
Related Documents