PDF-Vaccine immunology
Author : natalia-silvester | Published Date : 2015-08-11
Section 1 leading to conditions such as allergy autoimmunity or even premature death are being raised The objective of this chapter 19 Vaccine immunology 2 Chapter 2
Presentation Embed Code
Download Presentation
Download Presentation The PPT/PDF document "Vaccine immunology" is the property of its rightful owner. Permission is granted to download and print the materials on this website for personal, non-commercial use only, and to display it on your personal computer provided you do not modify the materials and that you retain all copyright notices contained in the materials. By downloading content from our website, you accept the terms of this agreement.
Vaccine immunology: Transcript
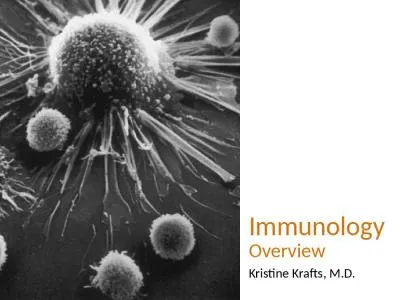
Section 1 leading to conditions such as allergy autoimmunity or even premature death are being raised The objective of this chapter 19 Vaccine immunology 2 Chapter 2 The conjugation of bacterial. . Allergology. . Pneumology. ENT. Pediatrics. US . AAAAI. Annual. 7-8000 . allergologists. . ATS. American . Thoracic. Society. Annual. 14000 . respiratory. . specialists. . AAOA. Annual. . ENT & . Kyle Murar. 5/5/2015. Period 4. What is Immunology?. Immunology is the study of the immune systems of all organisms.. Immunologists not only study the human immune system, but the immune systems organisms ranging from archaea bacteria to zebras.. 2014-2015 . Season: . What . you need to know. September 11, 2014. Cindy Schulte, RN, BSN. Kathy Sen, RN, BSN. Vaccine Preventable Disease Surveillance. Bureau of Immunization. New York State Department of Health (NYSDOH). of Asthma . through Biologics . Private Practice & St Michael’s Hospital. Lecturer, Division of Clinical Immunology & Allergy . Department of Medicine, University of Toronto. Jason Lee, MD, FRCPC. Presentation to: . Presented by:. Date:. Disclosure Statements. Neither the planners of this session nor I have any financial relationship with pharmaceutical companies, biomedical device manufacturers, or corporations whose products and services are related to the vaccines we discuss.. in situ. . – . elucidating the mechanisms of immune escape in adenovirus-induced . epidemic . keratoconjunctivitis. . (EKC). Makoto Yawata. 1,2. , . . Kevin John Selva. 2. , Jay Siak. 3. , Liu Yu Chi. Dr. Vibha Biotherapeutics group at Bristol Meyers Squibb. Previously, s he led the Predictive and Clinical Immunogenicity group Director of Preclinical Development at Merck Research Labs where s he Overview. Immunology Overview. Definitions. Cells. Lymphocytes . Antigen-presenting cells. Responses. The innate immune response. Capturing and displaying antigens. Cell-mediated immunity. Humoral immunity. Marrow Transplantation. Chapter 33. Tumor Immunology. Figure 33-01.. Embryonic primary germ layers. . (Redrawn from Larsen WJ:. Human embryology, . ed 3, Philadelphia, 2001, Churchill Livingstone.). THE GASTRINTESTINAL TRACT. T. HE GUT. Secretory Enterocytes. Absorptive Enterocytes. Tight Junction. (. claudins. , . occludins. and junctional adhesion molecules). Prevent . passive movement of hydrophilic. and Allergy Services. Daniel Clarke (Advanced Specialist Pharmacist). Amie Callow (Pharmacy Technician). University Hospital of North Midlands. Overview of UHNM Immunology and Allergy Service. UHNM Immunology and Allergy . ZOOLOGY Complement System 1 Development Team Paper Coordinator: Prof. Anju Srivastava Department of Zoology, University of Delhi Principal Investigator: Prof. Neeta Sehgal Head, Department o M. ajed. M. M.. L. Dr. . Mohanad. A. K.. * . What is Immunology. Historical background. Immune . system. Innate immunity. Immunity. . is defined as resistance to disease, specifically infectious disease and cancer. . Vasilis Pliasas. 1. , Zach Menne. 2. , Virginia Aida. 1. , Ji-Hang Yin. 1. , Peter J. Neasham. 1. , J. Fletcher North. 1. , Maria C. Naskou. 1. , Sheniqua Glover. 1. , Ioanna Skountzou. 2. , Constantinos S. .
Download Rules Of Document
"Vaccine immunology"The content belongs to its owner. You may download and print it for personal use, without modification, and keep all copyright notices. By downloading, you agree to these terms.
Related Documents