PPT-V25: the histone code 25. lecture WS 2019/20
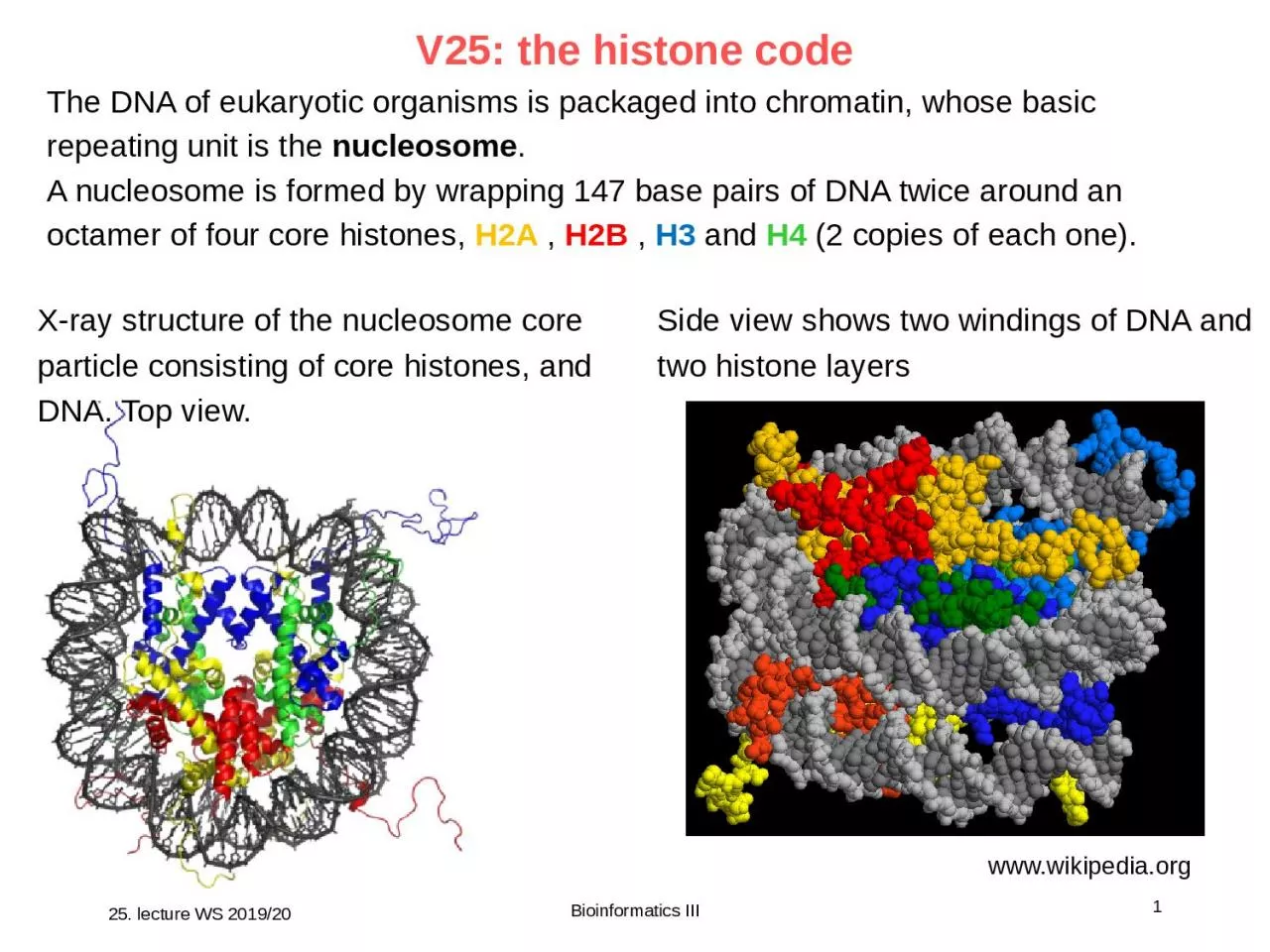
1 Bioinformatics III Xray structure of the nucleosome core particle consisting of core histones and DNA Top view wwwwikipediaorg Side view shows two windings of
Download Presentation
"V25: the histone code 25. lecture WS 2019/20" is the property of its rightful owner. Permission is granted to download and print materials on this website for personal, non-commercial use only, provided you retain all copyright notices. By downloading content from our website, you accept the terms of this agreement. Download
Presentation Transcript
Transcript not available.