PPT-Protein Structures Primary sequence
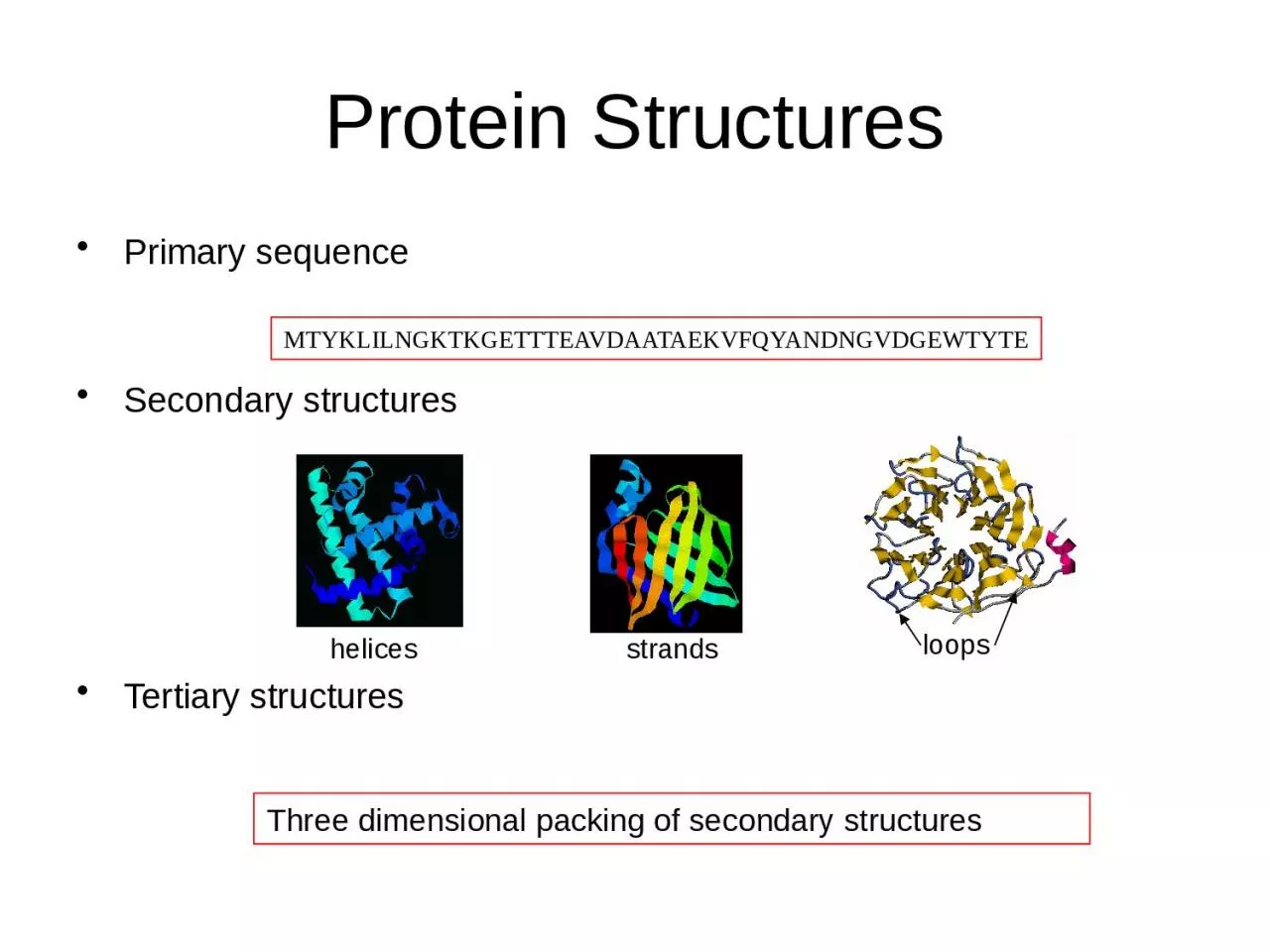
Secondary structures Tertiary structures MTYKLILNGKTKGETTTEAVDAATAEKVFQYANDNGVDGEWTYTE helices strands loops Three dimensional packing of secondary structures Protein
Download Presentation
"Protein Structures Primary sequence" is the property of its rightful owner. Permission is granted to download and print materials on this website for personal, non-commercial use only, provided you retain all copyright notices. By downloading content from our website, you accept the terms of this agreement. Download
Presentation Transcript
Transcript not available.